

赛福基因公开课《精神分裂症分子遗传学研究最新进展》
Part-1
嘉宾介绍
李军,医学博士,毕业于日本东京大学分子细胞生物学专攻神经细胞学分野。现就职于美国国立卫生研究院(NIH)下属国立神经疾病与脑中风研究所(NINDS),从事博士后研究。目前研究工作是自闭症、癫痫等神经精神系统疾病的分子发病机制。
Part-2
公开课环节

各位老师, 各位赛福基因的新老朋友,大家好,欢迎大家来到赛福基因大讲堂第二十四期的讲座。我是本期的主讲人,我叫李军, 目前在美国国立卫生研究院下属的神经疾病与脑中风研究所从事博士后研究。主要的研究方向是自闭症,癫痫,精神分裂症等神经精神系统疾病的分子发病机制。很高兴能够再次来到大讲堂,与各位朋友分享交流。这次和大家分享的主题是,精神分裂症分子遗传学研究的最新进展。下面,就开始我的报告。

首先在这张幻灯片上, 我列出了本次报告的内容提要,主要分为四个部分,首先介绍精神分裂症相关的背景知识, 包括精神分裂症的定义和诊断标准,其次介绍关于精神分裂症发病机制的几种假说,接下来从遗传学和表观遗传学两个角度,重点介绍近年来精神分裂症分子遗传学研究方面取得的一些进展,最后一部分,讨论一下精神分裂症研究面临的诸多挑战以及对于未来研究的展望。

在这张幻灯片上,我先向大家介绍几位著名的精神分裂症病人。左边的是梵高,荷兰著名画家,后印象主义先驱,代表画作有《星夜》《向日葵》等,他本人长期与精神分裂症斗争,出现有幻听、抑郁、自杀倾向等精神分裂症表现,左边的这两幅画是他自残割伤耳朵后的自画像。右边这位是草间弥生,日本当代著名的艺术家和作家,代表作有《无限镜屋》、《南瓜》等。她本人自幼年就罹患精神分裂症,经常出现幻听、幻视、并有自杀倾向,其大多数的时间在精神疗养院里休息、创作,游走于艺术家和精神病患者两种身份之间,是一位自愿住进精神疗养院的艺术家。下面这位是大名鼎鼎的约翰纳什教授,美国著名的数学家经济学家,因为博弈论方面的重要成就而获得1994年诺贝尔经济学奖。根据约翰纳什的生平以及与精神分裂症斗争的经历为基础而创作的电影《美丽心灵》,也广为人知。
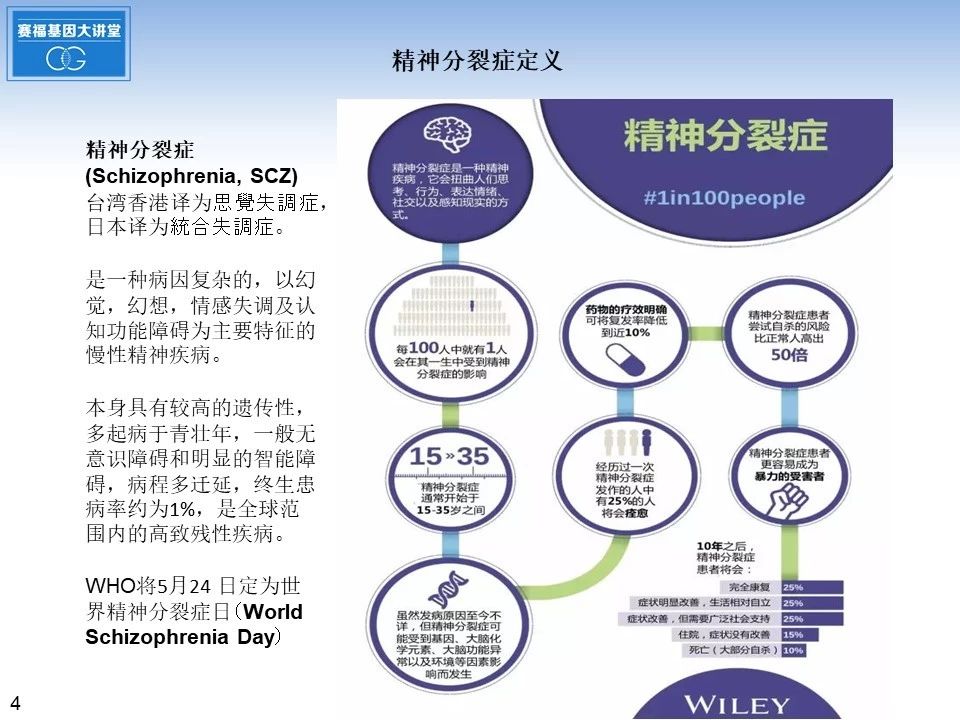
接下来我们切入正题,首先介绍一下精神分裂症的定义。精神分裂症,英文Schizophrenia, 为了避免污名化,台湾香港将其改译为思覺失調症,在日本译为統合失調症。精神分裂症,它是一种病因复杂的,以幻觉,幻想,情感失调及认知功能障碍为主要特征的慢性精神疾病。疾病本身具有较高的遗传性,多起病于青壮年15-35岁之间,一般无意识障碍和明显的智能障碍,病程多迁延,终生患病率约为1%,是全球范围内的高致残性疾病,严重影响患者本人的生活质量,也为家庭和社会造成沉重的精神心理压力和经济负担。世界卫生组织将每年的5月24 日定为世界精神分裂症日(WorldSchizophrenia Day)。

根据美国精神病学会颁布的精神障碍诊断及统计手册第五版DSM-5,确定精神分裂症的关键特征有如下五个:包括有妄想,幻觉,思维言语紊乱,异常的运动行为,还有阴性症状。

这张幻灯片介绍的是精神分裂症的诊断标准,根据《精神障碍诊断及统计手册》DSM-5上确定的,共有ABCDEF六项指标。A特征性症状:具有下列2项(或更多)症状,均应存在一月以上(如已经过有效治疗,病期可较短),其中的一项,必须是第1到3项症状之一;B社交或职业功能不良;C病期至少持续6月以上;D排除分裂情感性及心境障碍;E排除精神物质或一般躯体情况;F与广泛性发育障碍的关系,如有自闭症或其他广泛性发育障碍或别的儿童期起病的交流障碍的病史,除非出现明显的妄想或幻觉至少1月,才可另加精神分裂症的诊断。DSM-5 取消了精神分裂症的亚型分类。

关于精神分裂症的发病机制有以下五种假说,包括多巴胺假说,血清素5-羟色胺假说,谷氨酸假说,g-氨基丁酸假说和免疫炎症假说。
接下来的幻灯片我们重点介绍一下,精神分裂症分子遗传学研究的一些最新进展,将从遗传学和表观遗传学两方面具体展开。

目前研究表明,精神分裂症是由遗传因素和环境因素相互作用所导致的复杂性精神疾病。国内外关于精神分裂症的家系调查,双生子及寄养子研究均显示遗传因素在本病发生中期主导作用。分子遗传学连锁与关联分析研究显示,有多个与精神分裂症发生相关的染色体位点,具体可参看左下角的染色体示意图。目前我们已知的精神分裂症致病风险基因包括,精神分裂症1断裂基因(DISC1),Dysbindin基因(DTNBP1),儿茶酚氧位甲基转移酶基因(COMT),神经调节蛋白基因(NRG1,Neuregulin1), G蛋白信号调节基因(RGS4)及其他一些基因。
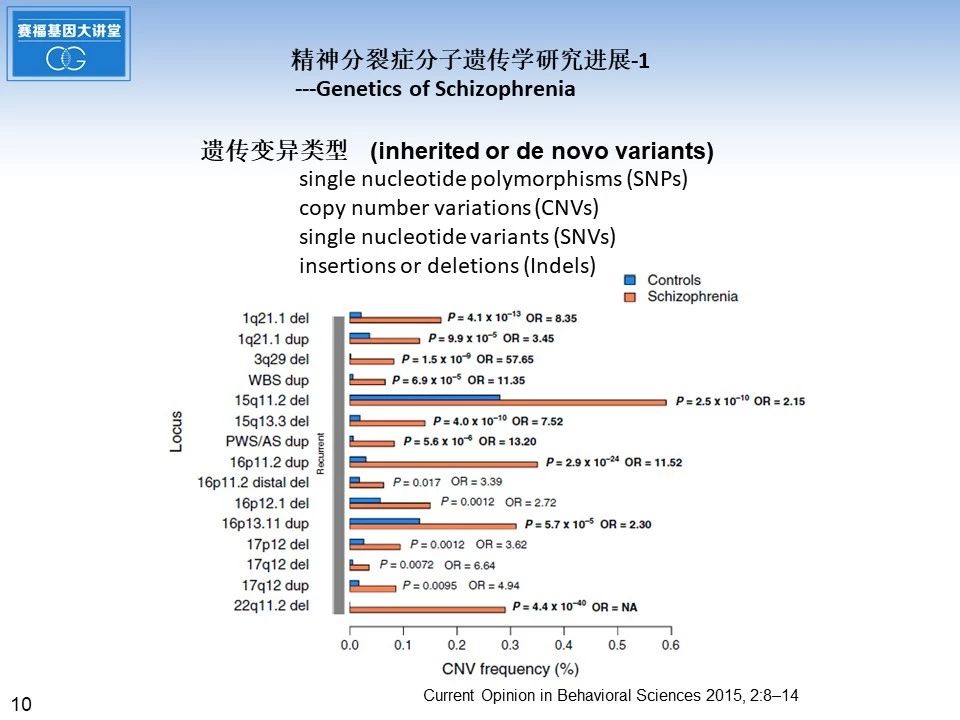
大量的研究发现,这些精神分裂症致病风险基因存在多种类型的变异,包括单核苷酸多态性SNP,拷贝数变异CNV, 单核苷酸变异SNV,以及插入和缺失突变,这些变异有些具有可遗传性,也有很多是新发突变de novo mutation。下边的柱状图是比较了在正常对照和精神分裂症病人之间,多个精神分裂症发病相关的染色体位点出现拷贝数变异的频率百分比。
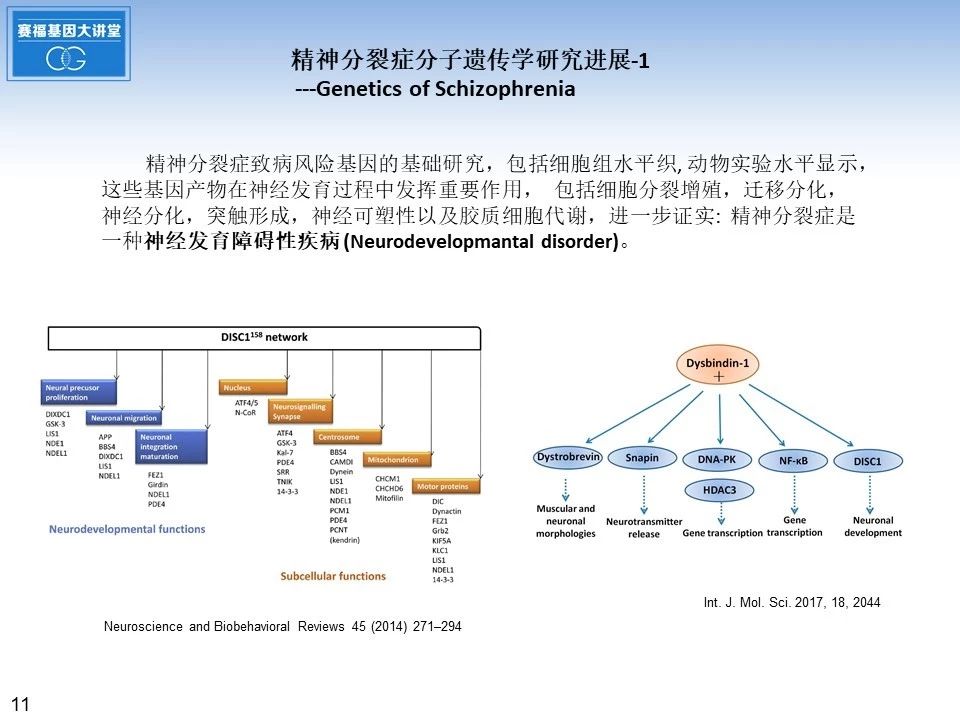
精神分裂症致病风险基因的基础研究,包括细胞组织水平, 动物实验水平显示,这些基因产物在神经发育过程中发挥重要作用,包括细胞分裂增殖,迁移分化,神经分化,突触形成,神经可塑性以及胶质细胞代谢,也进一步证实: 精神分裂症是一种神经发育障碍性疾病。下面是关于DISC1和Dysbindin-1两个经典的精神分裂症致病风险基因研究工作的总结示意图。DISC1基因位于1号染色体长臂1q42,Dysbindin-1基因位于6号染色体短臂6p22.3, 它们编码的蛋白质,可以和多种蛋白质分子相互作用,在神经发育过程中发挥重要作用。
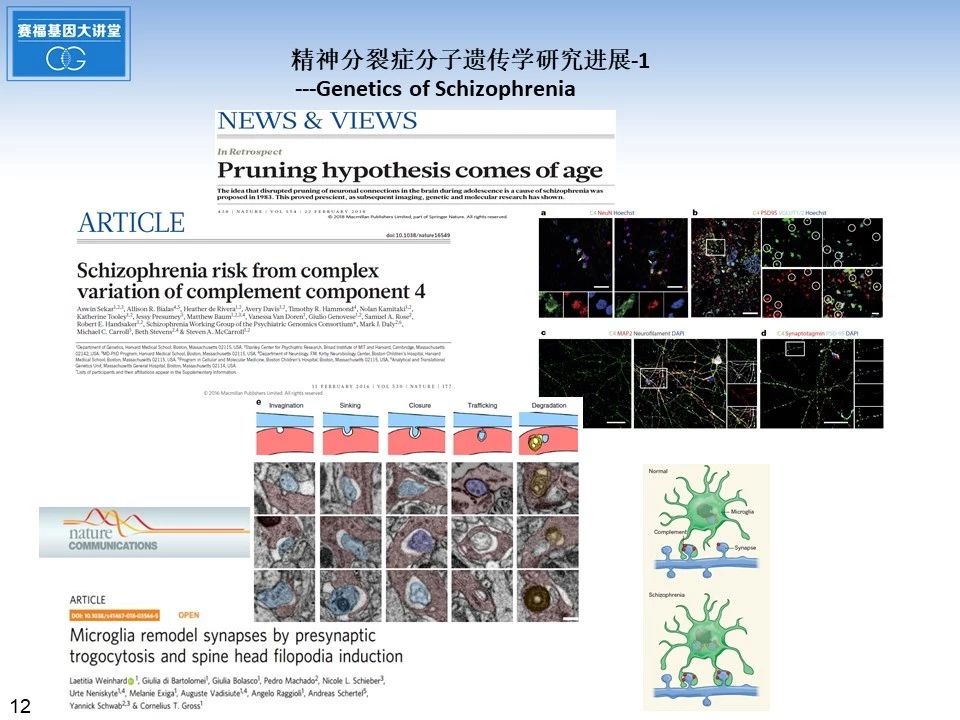
2016年二月《自然》杂志发表的研究论文显示,精神分裂症与主要组织相容性复合体(MHC)密切相关,补体成分C4表达在神经元的胞体和突触上,可以介导小胶质细胞参与的突触修剪过程,这为精神分裂症免疫炎症假说提供了证据支持。今年年初意大利的研究小组,尝试3种不同的成像系统, 结合电镜以及光片荧光显微镜, 首次在大脑中观察到了小胶质细胞“蚕食”突触的动态视频。这项研究发表在《自然通讯》上。《自然》杂志也发表评论性文章,欢呼突触修剪假说时代的到来。
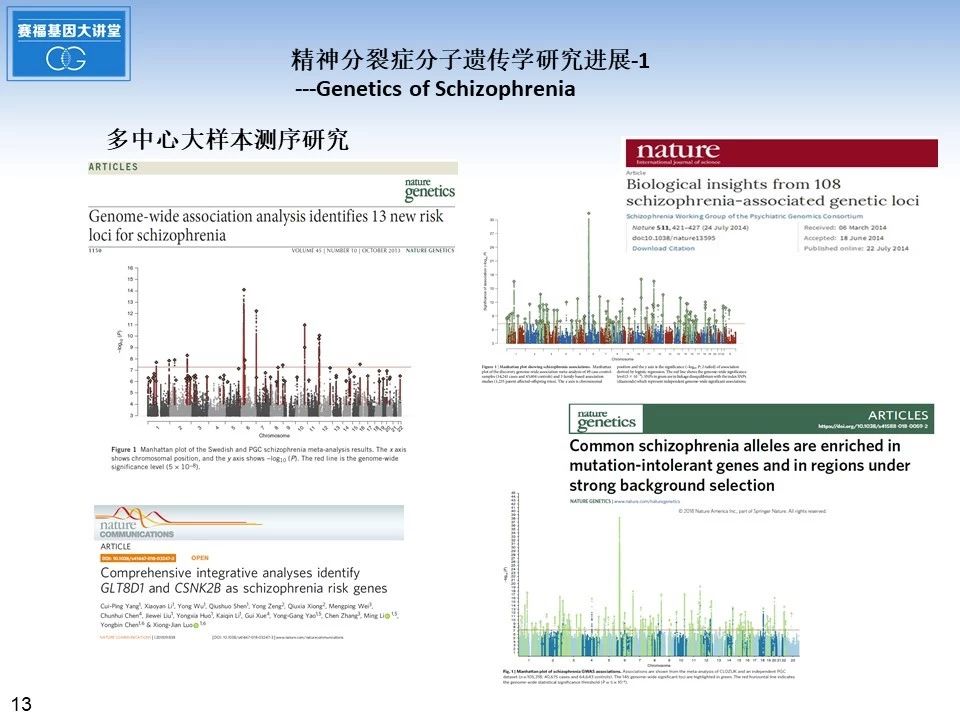
随着测序技术的发展和生物信息学分析方法的改进,近年来开展了多个多中心大样本的精神分裂症测序研究。2013年十月《自然遗传学》发表了瑞典人的精神分裂症基因组相关分析, 鉴定出13个新的精神分裂症风险基因位点。2014年七月《自然》杂志发表了精神疾病基因组协会(PsychiatricGenomics Consortium,PGC)的研究成果找到了108个精神分裂症相关的基因位点,发现多个位点都涉及神经传递,尤其是多巴胺相关的神经传递。随着被研究人群的数量不断增加,新发现的相关遗传位点的数量也在相应的增加。 2018年2月26日,《自然遗传学》杂志发表了来自英国卡迪夫大学、美国哈佛大学等全球几十家著名研究机构的重磅文章,在纳入超过一万名精神分裂症患者(对照组24542人,精神分裂症患者11260人)的大规模测序研究中,研究者们发现145个与精神分裂症相关的基因位点,其中50个为新发现的基因相关位点,首次显示常见的变异关联信号在受到强烈选择压力的基因中高度富集。同时期,中科院昆明动物研究所罗雄剑课题组在《自然通讯》发表论文, 使用整合基因组方法确定精神分裂症的候选致病基因, 研究显示ALMS1,GLT8D1和CSNK2B这三种基因在精神分裂症发病机制中的潜在作用。进一步研究显示GLT8D1和CSNK2B敲低促进神经干细胞的增殖和抑制分化能力,并改变神经元的形态学和突触传递。
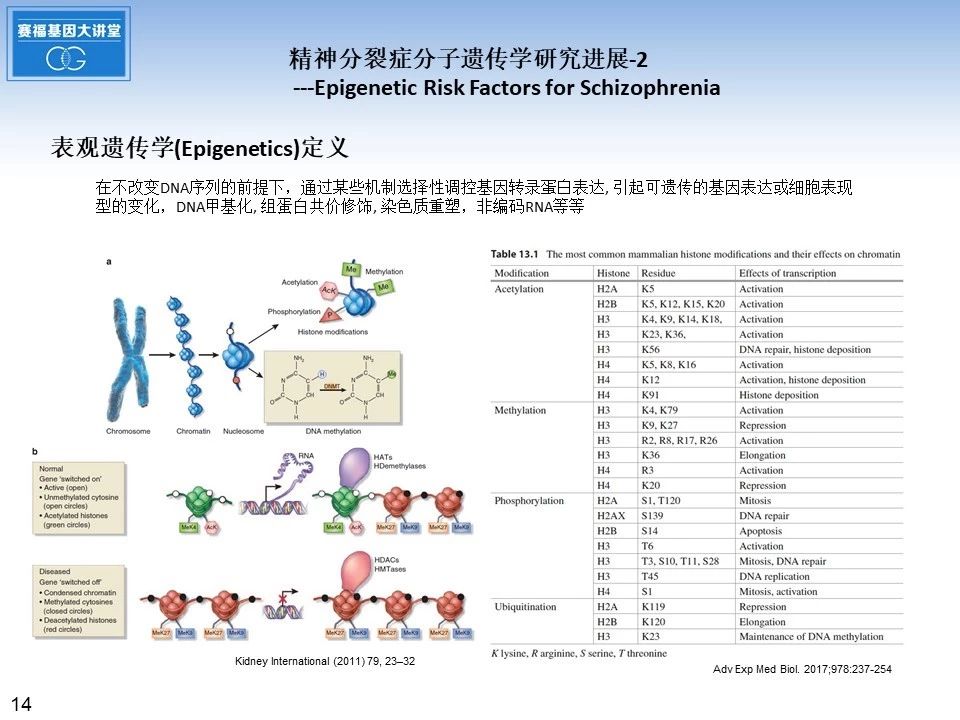
接下来的幻灯片,从表观遗传学角度,介绍近年来精神分裂症分子遗传学研究方面取得的一些进展. 首先介绍表观遗传学定义,在不改变DNA序列的前提下,通过某些机制选择性调控基因转录蛋白表达, 引起可遗传的基因表达或细胞表现型的变化,包括DNA甲基化, 组蛋白共价修饰, 染色质重塑,非编码RNA等等。
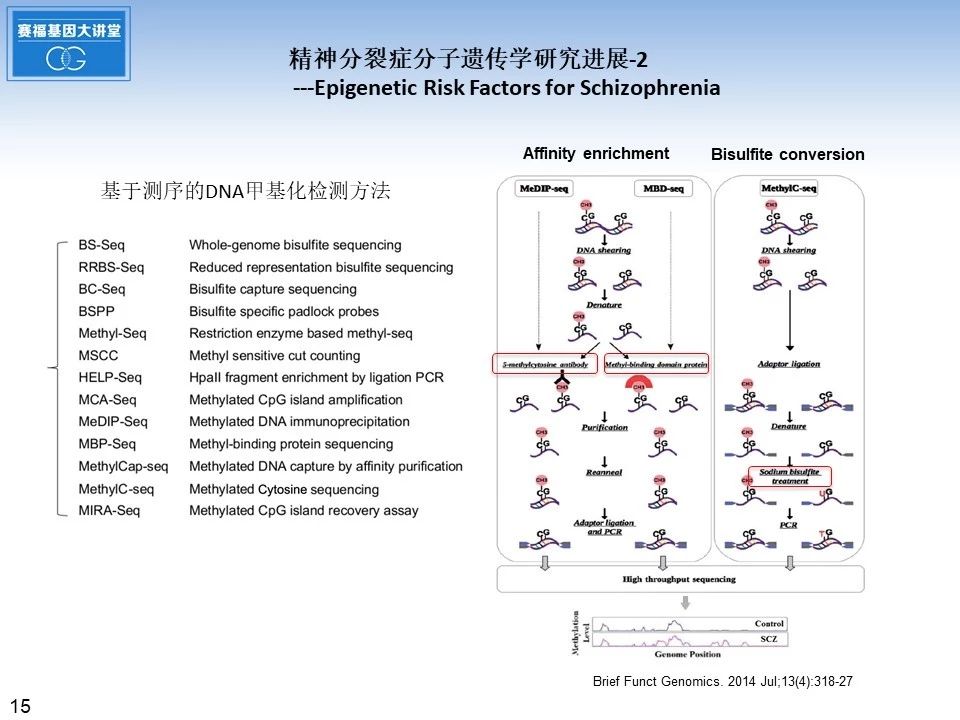
关于如何检测DNA甲基化水平,左侧我列举了一些基于测序的DNA甲基化检测方法,右边显示了几种常用的检测方法的流程图,首先将基因组DNA片段化处理,再利用5甲基胞嘧啶抗体或者甲基化结合蛋白进行亲和富集,或者是利用亚硫酸氢盐处理后,胞嘧啶可以转变尿嘧啶,而甲基化的胞嘧啶不发生变化,之后再用于高通量测序,进而比较DNA甲基化水平。

这张幻灯片上表格总结了近年来,利用精神分裂症病人不同区域的脑组织,利用表观基因组相关分析的方法,研究DNA甲基化水平,发现了很多表观遗传学标记物呈现出明显的组织特异性。
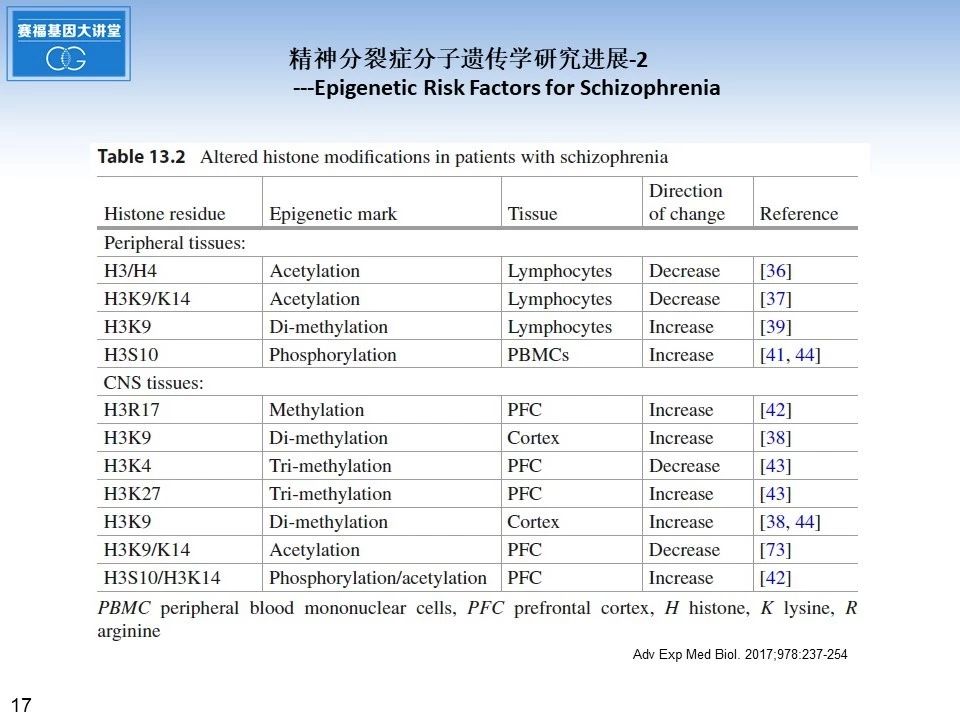
这张幻灯片上表格总结了近年来精神分裂症病人,利用淋巴细胞,外周血单个核细胞或者额叶皮质等不同样本,检测到的组蛋白修饰状态的不同变化。

最后一部分是,我们讨论一下精神分裂症研究面临的诸多挑战以及对于未来研究的展望。随着测序技术的不断发展,我们鉴定出越来越多的精神分裂症相关风险基因,这些基因,其中很多又与其他的精神疾病存在大量的重合,如何解析这些风险基因就成为一个巨大的挑战,分析他们在何时何处如何影响脑的发育和功能。再者,传统的基因敲除敲入模型已经不能充分模拟风险因子的分子病理学,我们需要构建新的基于特定变异类型的细胞动物模型,现在CRISPR/CAS9基因组编辑技术可以部分的满足研究的需要。再有就是,在细胞动物模型上,如何研究基因与环境的相互作用的机制,也变得越来越重要。
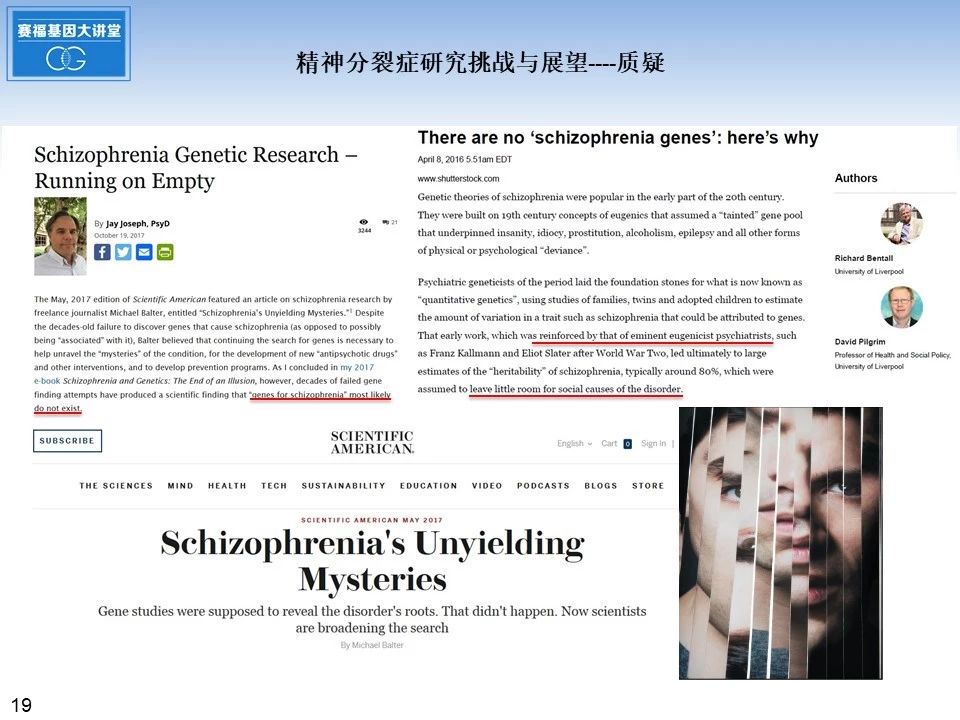
在精神分裂症DNA测序研究蓬勃开展的时候,同时也出现很多质疑的声音,有的认为可能并不存在所谓的精神分裂症基因,有的认为社会环境心理因素也影响精神分裂症的发病过程,然而长期以来没有得到重视,再有就是,精神分裂症的新药研发并没有因为基础研究的发展而出现重大突破。这些质疑的声音,很值得我们基础科研人员思考。

在2017年,世界生物精神病学会联盟WFSBP,发表了共识性论文,总结了当前精神分裂症遗传学研究的现状,也承认在基因环境相互作用和表观遗传学研究方面仍然有缺失,并且指出将来精神分裂症的研究要整合多种方法,包括基因组学,表观基因组学,转录组学,蛋白质组学,代谢组学等,探究精神分裂症的发病机制,指导新药研发,从而实现精神分裂症患者的个体化精准治疗。

在报告的最后,我要感谢大家的细心聆听,如果有不当之处 ,敬请大家批评指正。再次感谢赛福基因大讲堂全体同仁的通力协作,谢谢大家!最后,祝大家五一长假快乐!
Part-3
问答环节
观众a:
李军老师您好,近几年来,测序技术极大促进了精神分裂症的基础研究,我想问一下现在在分子诊断方面是否有比较成熟的生物标记物Biomarkers或者用于基因检测的Panel?
李军老师:
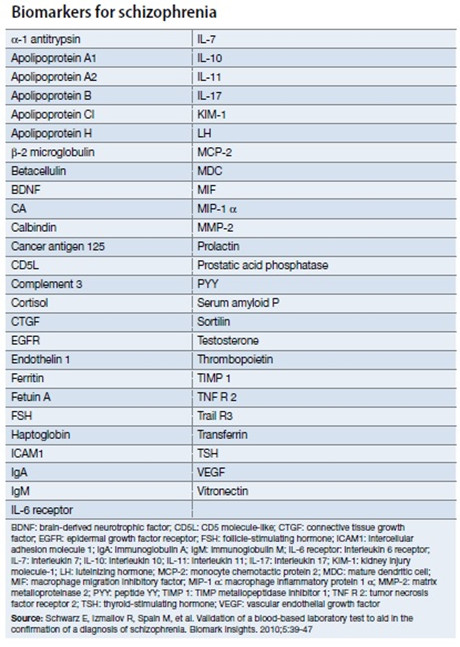
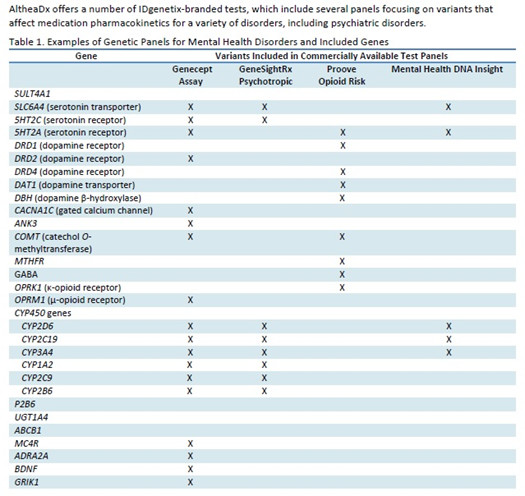
如何寻找比较可靠的生物标记物,对于精神分裂症的提早预防提早治疗,确实非常重要,这也是精神分裂症基础研究的一个热点方向。在2010年发表过一篇论文,声称开发出了一个含有51个标记物的精神分裂症的血液检查。但是由于精神分裂症病因的复杂性,目前并没有在临床广泛应用。关于你所说的精神分裂症基因检测的Panel,目前还没有针对的检测服务项目。但是,在美国有基因检测机构推出Mental Health Disorders的基因检测Panel,其中包含有多个精神分裂症风险基因。具体可以参见下列示意表格。
观众b:
请问李老师,现在非编码RNA在精神分裂症研究中的进展如何?
李军老师:
这位老师问的是非编码RNA在精神分裂症研究中的一些进展,这是一个非常好的问题。非编码RNA的研究本身就是生物医学研究的一个热门方向,也是表观遗传学研究的一个重要的分支,尤其在研究肿瘤发病机制方面。非编码RNA在神经系统发育过程也发挥重要的作用,如调节神经分化,突触形成,突触传递及突触可塑性等等。由于时间和篇幅所限,在这里没做介绍。如果您感兴趣的话,我可以推荐您两篇综述,值得一读。具体信息详见下图。


观众c:
感谢李老师的精彩报告,现在关于精神分裂症的表观遗传学研究,DNA甲基化可以用测序的方法检测,组蛋白的不同类型的修饰,也可以测序检出吗?还是说用其他的方法?
李军老师:
您问了一个非常细致的技术问题,DNA甲基化可以用测序的方法检测,但是组蛋白的不同类型的修饰,是不能用测序的方法检出的。目前检测组蛋白的不同修饰类型,主要还是应用特异性的抗体进行Western blot检测,或者更精确的方法就是进行组蛋白的质谱分析。
观众d:
李老师您好,在讲座中您提到将来要加强精神分裂症在基因与环境相互作用方面的机制研究,不知道都用些什么研究方法啊?
李军老师:
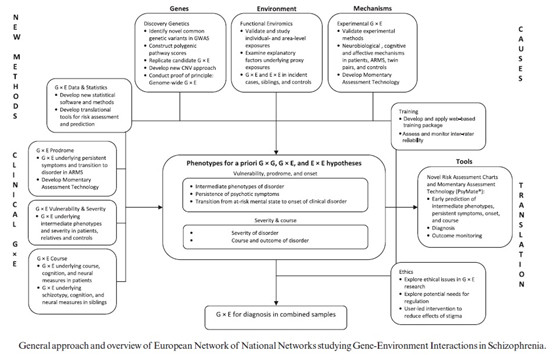
精神分裂症的发生与多种社会环境心理因素相关,包括文化,职业和社会阶层,移民,孕期饥饿,社会隔离及突发应激事件等等。我们可以开展这些特定人群的分子遗传学检测,包括遗传学和表观遗传学等多个层面,应该可以发现更多的更具有特异性的基因突变位点。再有,对于已有的和新构建的精神分裂症风险基因的敲除敲入动物模型,再外加特定环境因素后,比如,孕期饥饿,社会隔离或突发应激事件等,进一步观察对于精神分裂症发病的影响以及在分子遗传学水平的变化。最近,欧洲精神分裂症基因环境相互作用研究网络(EU-GEI)发布了一个工作框架图(见下图),相关文章,很值得一读。

观众e:
请教下李老师,现在自闭症的分子遗传方面的研究现状,谢谢!
李军老师:
这位老师问的是自闭症分子遗传学方面的研究现状。其实自闭症作为一种神经精神系统疾病,与精神分裂症的研究方法手段上比较相似。最近关于自闭症测序方面有许多的文章发表。利用不同的高通量测序方法,对不同的人群,或者对不同组织以及不同细胞组分进行特定的NGS测序,包括WGS,WES或者其他的测序方法。具体内容可以点击此处查看。